ژنها (مثلاً: jeenz ) نقش مهمی در تعیین ویژگیهای فیزیکی - ظاهر ما - و بسیاری چیزهای دیگر در مورد ما دارند. آنها حاوی اطلاعاتی هستند که باعث می شود شما چه کسی هستید و چه شکلی هستید: موهای مجعد یا صاف، پاهای بلند یا کوتاه، حتی اینکه چگونه ممکن است لبخند بزنید یا بخندید. بسیاری از این چیزها از یک نسل به نسل دیگر در یک خانواده توسط ژن ها منتقل می شود.
در پایان این درس، شما می دانید
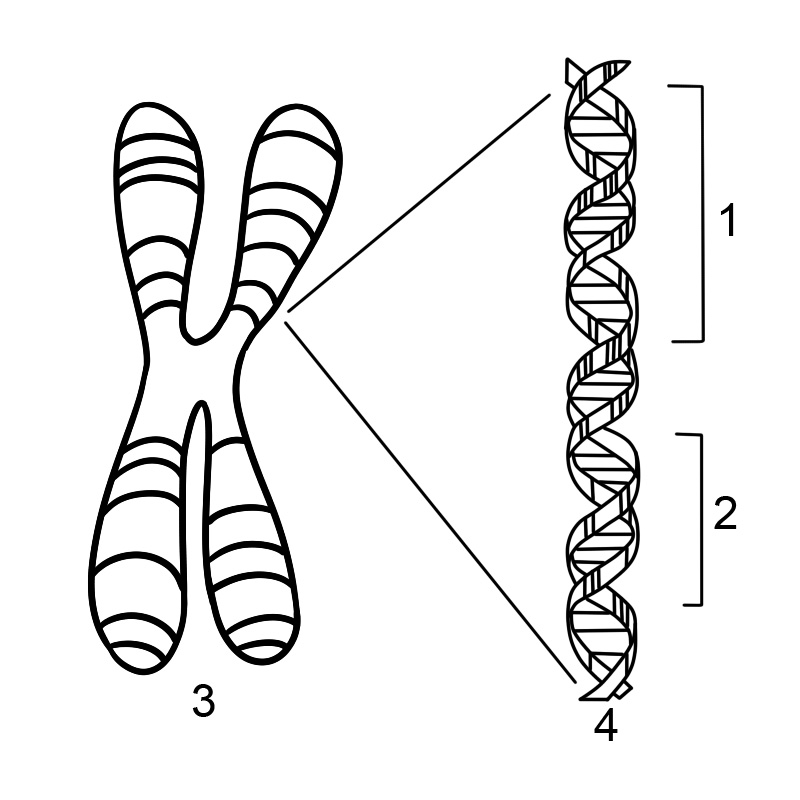
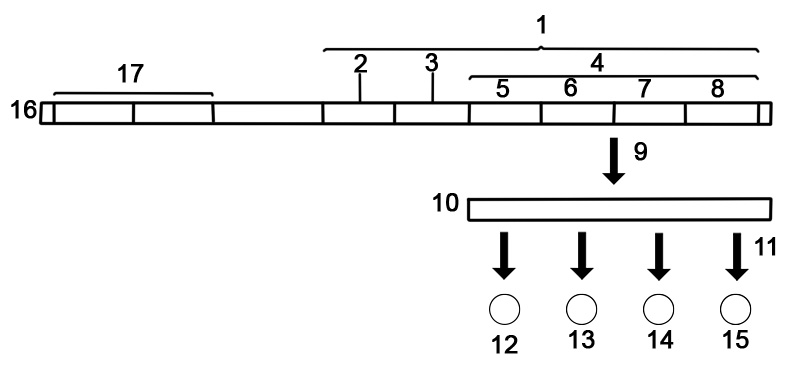
یک ژن دنباله ای از نوکلئوتیدها در DNA یا RNA است که سنتز یک محصول ژن، RNA یا پروتئین را رمزگذاری می کند. یک کروموزوم از یک رشته DNA طولانی که دارای ژن های زیادی است تشکیل شده است. یک کروموزوم انسانی می تواند تقریباً 500 میلیون جفت باز DNA با هزاران ژن داشته باشد.
در زیستشناسی، یک ژن به دنبالهای از نوکلئوتیدها در RNA یا DNA اطلاق میشود که برای مولکولی که عملکردی دارد، کد میکند. در طول بیان یک ژن، DNA ابتدا در RNA کپی می شود. RNA می تواند مستقیماً کاربردی باشد یا می تواند الگوی واسطه ای برای پروتئینی باشد که یک عملکرد را انجام می دهد. انتقال ژن ها به فرزندان یک ارگانیسم اساس وراثت صفات فنوتیپی را تشکیل می دهد. این ژن ها توالی های DNA مختلفی را می سازند که به نام ژنوتیپ شناخته می شوند. ژنوتیپ ها به همراه عوامل محیطی و رشدی تعیین می کنند که فنوتیپ چه خواهد بود. بیشتر صفات بیولوژیکی تحت تأثیر پلی ژن ها (بسیاری از ژن های مختلف) و برهمکنش های ژن-محیط هستند. برخی از صفات ژنتیکی مانند رنگ چشم قابل مشاهده هستند و برخی دیگر مانند گروه خونی نیستند.
این امکان وجود دارد که ژن ها در توالی خود جهش پیدا کنند. این منجر به انواع مختلفی به نام آلل در جمعیت می شود. این آلل ها نسخه های کمی متفاوت از یک پروتئین را رمزگذاری می کنند که باعث ایجاد صفات فنوتیپی می شود. ژن ها در نتیجه انتخاب طبیعی یا بقای مناسب ترین و رانش ژنتیکی آلل ها تکامل می یابند.
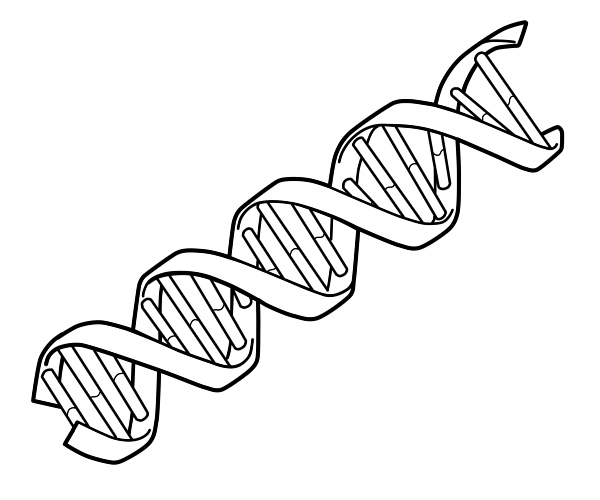
در اینجا تصویری است که رشته دو مارپیچ DNA را نشان می دهد.
بیشتر موجودات زنده ژن های خود را در رشته های طولانی DNA رمزگذاری می کنند. DNA مخفف دئوکسی ریبونوکلئیک اسید است. DNA از زنجیره ای تشکیل شده است که از چهار نوع زیر واحد نوکلئوتیدی تشکیل شده است که هر کدام از یک قند پنج کربنه (2-دئوکسی ریبوز)، یک گروه فسفات و یکی از چهار باز آدنین، تیمین، سیتوزین و گوانین تشکیل شده است.
دو زنجیره DNA به دور یکدیگر می پیچند و یک مارپیچ دوگانه DNA را تشکیل می دهند که بازها به سمت داخل و بازهای آدنین به تیمین و گوانین به سیتوزین جفت می شوند. ویژگی جفت شدن بازها به این دلیل اتفاق می افتد که آدنین و تیمین برای تشکیل دو پیوند هیدروژنی در یک راستا قرار می گیرند. سیتوزین و گوانین از طرف دیگر سه پیوند هیدروژنی تشکیل می دهند. دو رشته در یک مارپیچ دوتایی باید مکمل یکدیگر باشند و توالی پایه آنها مطابقت داشته باشند به طوری که آدنین های یک رشته با تیمین های رشته دیگر جفت شوند و غیره.
بیان ژنهای کدگذاری شده در DNA با رونویسی ژن به RNA، نوع دوم اسید نوکلئیک که مونومرهای آن از ریبوز قند ساخته شدهاند، به جای دیاکسی ریبوز مانند DNA، آغاز میشود. RNA همچنین حاوی اوراسیل پایه به جای تیمین است. مولکول های RNA تک رشته ای هستند و نسبت به DNA پایداری کمتری دارند. ژن هایی که پروتئین ها را کد می کنند از یک سری توالی سه نوکلئوتیدی به نام کدون ساخته شده اند. کد ژنتیکی مطابقت را در طول ترجمه پروتئین بین کدون ها و اسیدهای آمینه مشخص می کند. کد ژنتیکی برای همه موجودات شناخته شده تقریبا یکسان است.
ساختار یک ژن متشکل از عناصر بسیاری است که توالی کدکننده پروتئین واقعی اغلب تنها بخش کوچکی از آنهاست. اینها شامل نواحی DNA که رونویسی نشده اند و همچنین مناطق ترجمه نشده RNA هستند.
ژن ها حاوی یک توالی تنظیمی هستند که زمان و مکان بیان برای ناحیه کد کننده پروتئین را کنترل می کند. اول، ژن ها به یک توالی پروموتر نیاز دارند. پروموتر توسط فاکتورهای رونویسی که جذب و به اتصال RNA پلیمراز به منطقه برای شروع رونویسی کمک می کنند، شناسایی و متصل می شود. تشخیص معمولاً به عنوان یک توالی اجماع مانند جعبه TATA رخ می دهد. یک ژن میتواند بیش از یک پروموتر داشته باشد، در نتیجه RNAهای پیامرسان (mRNA) ایجاد میشود که در میزان گسترش آنها در انتهای 5' متفاوت است. ژنهای بسیار رونویسیشده دارای توالیهای پروموتر «قوی» هستند و ژنهای دیگر دارای پروموترهای «ضعیف» هستند که ارتباط ضعیفی با عوامل رونویسی ایجاد میکنند و رونویسی را کمتر آغاز میکنند. شناسایی نواحی پروموتر یوکاریوتی بسیار پیچیده تر و دشوارتر از پروموترهای پروکاریوتی است.
تقویتکنندهها رونویسی را با اتصال به پروتئین فعالکننده افزایش میدهند که سپس به جذب RNA پلیمراز به پروموتر کمک میکند. برعکس، خفه کن ها به پروتئین های سرکوب کننده متصل می شوند و DNA را برای RNA پلیمراز کمتر در دسترس قرار می دهند. pre-mRNA رونویسی شده شامل مناطق ترجمه نشده در دو انتها است که حاوی یک محل اتصال ریبوزوم، پایان دهنده و کدون های شروع و توقف است. علاوه بر این، بیشتر یوکاریوت ها حاوی اینترون های ترجمه نشده هستند که قبل از ترجمه اگزون ها حذف می شوند. توالیهای انتهای اینترونها ، مکانهای اتصال را برای تولید mRNA بالغ نهایی که پروتئین یا محصول RNA را کد میکند، دیکته میکنند.
در زیر ساختار یک ژن کد کننده پروتئین یوکاریوتی آمده است.
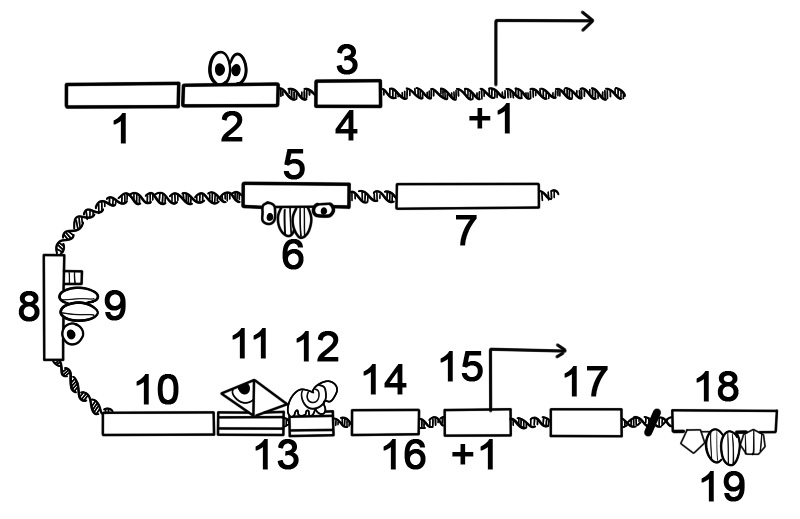
بسیاری از ژنهای پروکاریوتی به اپرونها، با توالیهای کدکننده پروتئین متعددی که بهصورت یک واحد رونویسی میشوند، سازماندهی میشوند. ژن های موجود در اپرون به صورت mRNA پیوسته رونویسی می شوند که به آن mRNA پلی سیسترونیک می گویند. در این زمینه، اصطلاح سیسترون معادل ژن است. رونویسی mRNA یک اپرون اغلب توسط یک رپرسور کنترل می شود که بسته به وجود متابولیت های خاص می تواند در حالت فعال یا غیرفعال رخ دهد. هنگامی که فعال است، رپرسور به یک توالی DNA در ابتدای اپرون، به نام ناحیه عملگر، متصل می شود و رونویسی اپرون را سرکوب می کند. هنگامی که رپرسور غیرفعال است، رونویسی اپرون ممکن است رخ دهد. محصولات ژنهای اپرون معمولاً عملکردهای مرتبطی دارند و در یک شبکه نظارتی درگیر هستند.
در زیر ساختار یک اپرون پروکاریوتی از ژن های کد کننده پروتئین آورده شده است.