遺伝子 (たとえばjeenz ) は、身体的特徴 (私たちの外見) や、私たちに関する他の多くの要素を決定する上で重要な役割を果たします。それらは、あなたが誰であるか、どのように見えるかを示す情報を運びます: 巻き毛またはストレートヘア、長い脚または短い脚、さらには笑顔や笑い方まで.これらの多くは、遺伝子によって家族内で世代から世代へと受け継がれます。
このレッスンの終わりまでに、あなたは知っているでしょう
遺伝子は、RNA またはタンパク質のいずれかの遺伝子産物の合成をコードする DNA または RNA のヌクレオチドのシーケンスです。染色体は、多くの遺伝子を持つ長い DNA 鎖で構成されています。ヒトの染色体には、数千の遺伝子を持つ約 5 億塩基対の DNA があります。
生物学では、遺伝子とは、機能を持つ分子をコードする RNA または DNA 内のヌクレオチドの配列を指します。遺伝子の発現中、DNA は最初に RNA にコピーされます。 RNA は直接機能する場合もあれば、機能を実行するタンパク質の中間テンプレートになる場合もあります。生物の子孫への遺伝子の伝達は、表現型形質の遺伝の基礎を形成します。これらの遺伝子は、遺伝子型として知られるさまざまな DNA 配列を構成します。遺伝子型と環境要因および発生要因によって、表現型がどのようなものになるかが決まります。ほとんどの生物学的形質は、ポリジーン (多くの異なる遺伝子) および遺伝子と環境の相互作用によって影響を受けます。目の色のように目に見える遺伝的形質もあれば、血液型のように見えないものもあります.
遺伝子がその配列に突然変異を獲得することは可能です。これにより、集団内に対立遺伝子と呼ばれるさまざまなバリアントが生じます。これらの対立遺伝子は、表現型の形質を引き起こすわずかに異なるバージョンのタンパク質をコードしています。遺伝子は、自然選択または適者生存および対立遺伝子の遺伝的浮動の結果として進化します。
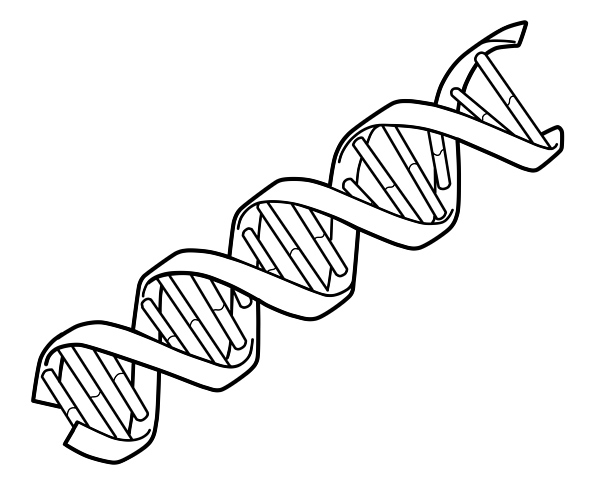
これは、DNA の二重らせん鎖を描いた図です。
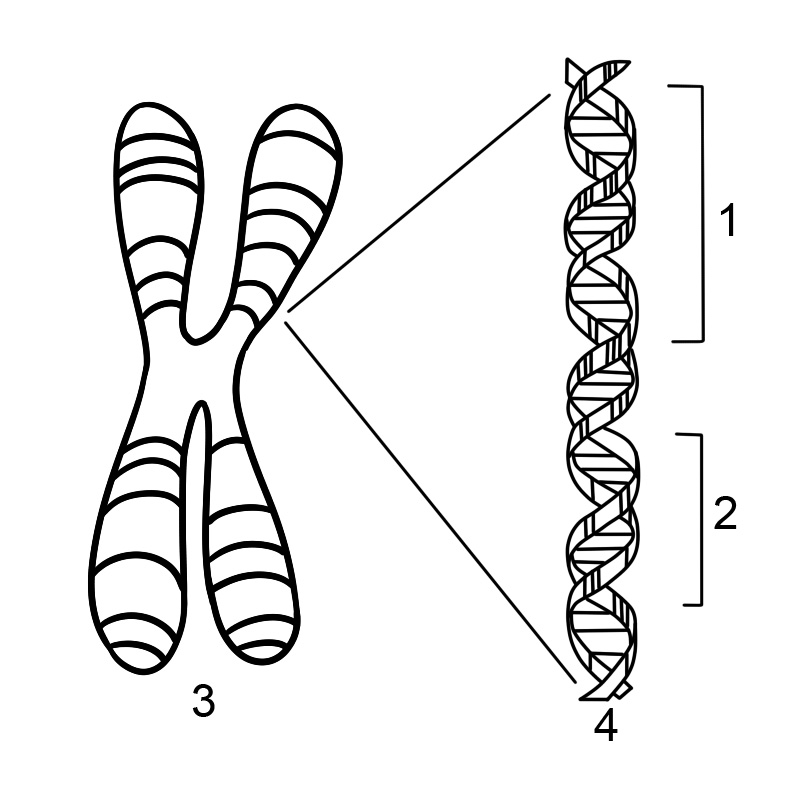
ほとんどの生物は、その遺伝子を長い DNA 鎖にエンコードします。 DNAはデオキシリボ核酸の略です。 DNA は 4 種類のヌクレオチド サブユニットからなる鎖で構成されており、各サブユニットは 5 炭素糖 (2-デオキシリボース)、リン酸基、および 4 つの塩基 (アデニン、チミン、シトシン、およびグアニン) のいずれかで構成されています。
2 本の DNA 鎖が互いにねじれて DNA 二重らせんを形成し、塩基が内側を向き、アデニン塩基がチミンに、グアニンがシトシンに対合します。塩基対形成の特異性は、アデニンとチミンが整列して 2 つの水素結合を形成するために起こります。一方、シトシンとグアニンは 3 つの水素結合を形成します。二重らせんの 2 本の鎖は、一方の鎖のアデニンがもう一方の鎖のチミンと対になるように、塩基配列が一致して相補的でなければなりません。
DNA にコードされた遺伝子の発現は、遺伝子を RNA に転写することから始まります。RNA は、DNA のようにデオキシリボースではなく糖リボースでモノマーが作られている 2 番目のタイプの核酸です。 RNA には、チミンの代わりに塩基ウラシルも含まれています。 RNA 分子は一本鎖であり、DNA ほど安定ではありません。タンパク質をコードする遺伝子は、コドンとして知られる一連の 3 つのヌクレオチド配列で構成されています。遺伝暗号は、コドンとアミノ酸の間のタンパク質翻訳中の対応を指定します。遺伝暗号は、既知のすべての生物でほぼ同じです。
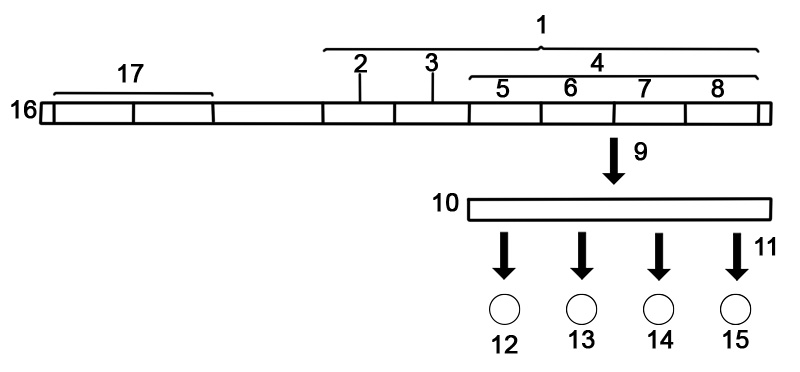
遺伝子の構造は多くの要素で構成されており、実際のタンパク質コード配列は多くの場合ほんの一部です。これらには、転写されない DNA 領域や RNA の非翻訳領域が含まれます。
遺伝子には、タンパク質コード領域の発現がいつどこで発生するかを制御する調節配列が含まれています。まず、遺伝子にはプロモーター配列が必要です。プロモーターは、RNAポリメラーゼがその領域に結合して転写を開始するのを助ける転写因子によって認識および結合されます。認識は通常、TATA ボックスのようなコンセンサス シーケンスとして発生します。遺伝子は複数のプロモーターを持つことができ、その結果、5' 末端での伸長の程度が異なるメッセンジャー RNA (mRNA) が生成されます。高度に転写された遺伝子には「強い」プロモーター配列があり、他の遺伝子には「弱い」プロモーターがあり、転写因子と弱い結合を形成し、転写を開始する頻度が低くなります。真核生物のプロモーター領域は、原核生物のプロモーターよりもはるかに複雑で、同定が困難です。
エンハンサーは、 RNAポリメラーゼをプロモーターに動員するのに役立つ活性化タンパク質に結合することにより、転写を増加させます。逆に、サイレンサーはリプレッサータンパク質に結合し、DNA を RNA ポリメラーゼに利用しにくくします。転写されたプレmRNAには、リボソーム結合部位、ターミネーター、開始および停止コドンを含む非翻訳領域が両端に含まれています。さらに、ほとんどの真核生物には、エクソンが翻訳される前に除去される未翻訳のイントロンが含まれています。イントロンの末端にある配列は、スプライス部位を指示して、タンパク質または RNA 産物をコードする最終的な成熟 mRNA を生成します。
以下は、真核生物のタンパク質コード遺伝子の構造です。
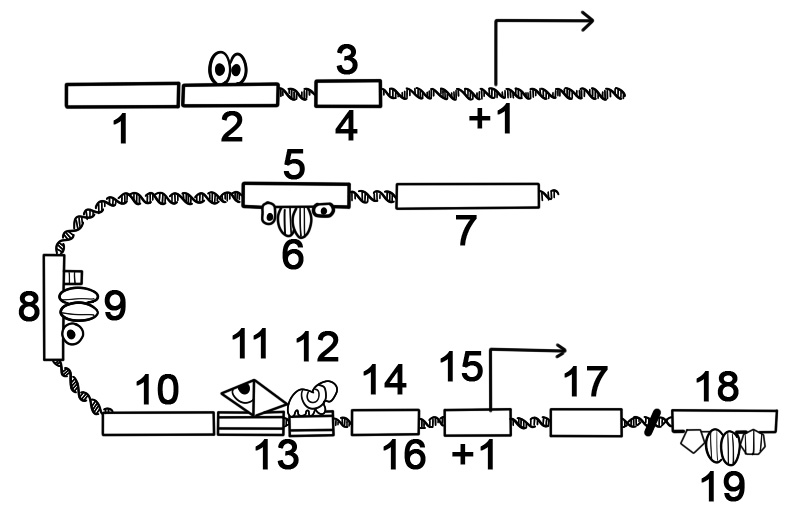
多くの原核生物の遺伝子は、ユニットとして転写される複数のタンパク質コード配列を持つオペロンに編成されています。オペロンの遺伝子は、ポリシストロン性 mRNA と呼ばれる連続した mRNA として転写されます。この文脈では、シストロンという用語は遺伝子に相当します。オペロンの mRNA の転写は、多くの場合、特定の代謝産物の存在に応じて活性または非活性の状態で発生するリプレッサーによって制御されます。リプレッサーが活性化すると、オペレーター領域と呼ばれるオペロンの先頭にある DNA 配列に結合し、オペロンの転写を抑制します。リプレッサーが不活性であるとき、オペロンの転写が起こります。オペロン遺伝子の産物は通常、関連する機能を持ち、同じ調節ネットワークに関与しています。
以下は、タンパク質をコードする遺伝子の原核オペロンの構造です。