Gener (säg: jeenz ) spelar en viktig roll för att bestämma fysiska egenskaper - hur vi ser ut - och massor av andra saker om oss. De bär information som gör dig till den du är och hur du ser ut: lockigt eller rakt hår, långa eller korta ben, till och med hur du kan le eller skratta. Många av dessa saker överförs från en generation till nästa i en familj genom gener.
I slutet av den här lektionen kommer du att veta
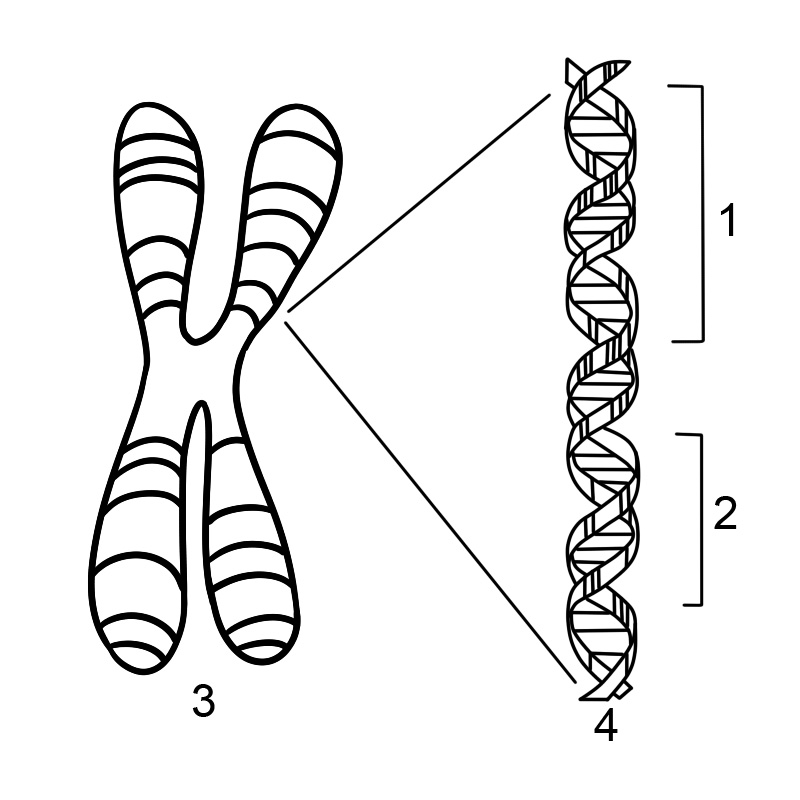
En gen är en sekvens av nukleotider i DNA eller RNA som kodar för syntesen av en genprodukt, antingen RNA eller protein. En kromosom är uppbyggd av en lång DNA-sträng som har många gener. En mänsklig kromosom kan ha cirka 500 miljoner baspar DNA med tusentals gener.
Inom biologi avser en gen en sekvens av nukleotider i RNA eller DNA som kodar för molekylen som har en funktion. Under uttrycket av en gen kopieras DNA först till RNA. RNA:t kan vara direkt funktionellt eller det kan vara den mellanliggande mallen för ett protein som utför en funktion. Överföringen av gener till en organisms avkomma utgör grunden för nedärvningen av de fenotypiska egenskaperna. Dessa gener utgör olika DNA-sekvenser som kallas genotyper . Genotyper tillsammans med miljö- och utvecklingsfaktorer avgör vad fenotypen blir. De flesta biologiska egenskaper påverkas av polygener (många olika gener) och gen-miljö-interaktioner. Vissa genetiska egenskaper är synliga som ögonfärg , och vissa är inte som blodgrupp.
Det är möjligt för gener att förvärva mutationer i sin sekvens. Detta leder till olika varianter som kallas alleler i befolkningen. Dessa alleler kodar för lite olika versioner av ett protein som orsakar fenotypiska egenskaper. Gener utvecklas som ett resultat av naturligt urval eller överlevnad av de starkaste och genetisk drift av allelerna.
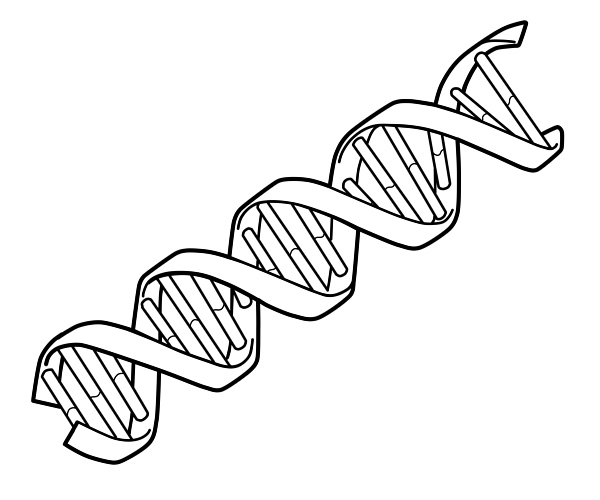
Här är en illustration som visar dubbelhelix-strängen av DNA.
De flesta organismer kodar för sina gener i långa DNA-strängar. DNA står för deoxiribonukleinsyra. DNA består av en kedja som består av fyra typer av nukleotidsubenheter, som var och en består av ett socker med fem kolatomer (2-deoxiribos), en fosfatgrupp och en av de fyra baserna adenin, tymin, cytosin och guanin.
Två DNA-kedjor vrider sig runt varandra för att bilda en DNA-dubbelhelix med baserna pekande inåt och adeninbaspar till tymin och guanin till cytosin. Specificiteten för basparning sker på grund av att adenin och tymin anpassas till två vätebindningar. Cytosin och guanin bildar å andra sidan tre vätebindningar. De två strängarna i en dubbelhelix måste vara komplementära med deras bassekvensmatchning så att adeninerna i en sträng paras ihop med tyminerna i den andra strängen, och så vidare.
Uttrycket av gener som kodas i DNA börjar med att transkribera genen till RNA, en andra typ av nukleinsyra vars monomerer är gjorda av sockerribosen snarare än deoxiribos som i DNA. RNA innehåller också basen uracil istället för tymin. RNA-molekyler är enkelsträngade och de är mindre stabila än DNA. Gener som kodar för proteiner är uppbyggda av en serie av tre-nukleotidsekvenser som kallas kodon. Den genetiska koden specificerar överensstämmelsen under proteintranslation mellan kodon och aminosyror. Den genetiska koden är nästan densamma för alla kända organismer.
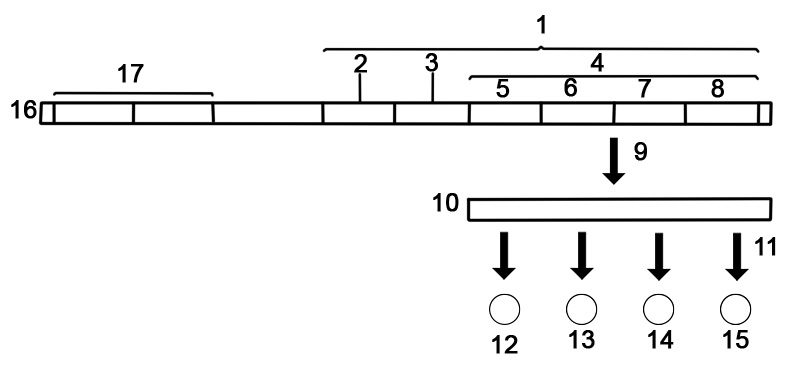
En gens struktur består av många element av vilka själva proteinkodningssekvensen ofta bara är en liten del. Dessa inkluderar DNA-regioner som inte transkriberas såväl som otranslaterade regioner av RNA:t.
Gener innehåller en regulatorisk sekvens som styr när och var expression sker för den proteinkodande regionen. För det första kräver gener en promotorsekvens . Promotorn känns igen och binds av transkriptionsfaktorer som rekryterar och hjälper RNA-polymeras att binda till regionen för att initiera transkription. Igenkänningen sker vanligtvis som en konsensussekvens som TATA-boxen. En gen kan ha mer än en promotor, vilket resulterar i budbärar-RNA (mRNA) som skiljer sig åt i hur långt de sträcker sig i 5'-änden. Högt transkriberade gener har "starka" promotorsekvenser och andra gener har "svaga" promotorer som bildar svaga associationer med transkriptionsfaktorer och initierar transkription mer sällan. Eukaryota promotorregioner är mycket mer komplexa och svåra att identifiera än prokaryota promotorer.
Enhancers ökar transkriptionen genom att binda ett aktivatorprotein som sedan hjälper till att rekrytera RNA-polymeraset till promotorn; omvänt binder ljuddämpare repressorproteiner och gör DNA:t mindre tillgängligt för RNA-polymeras. Det transkriberade pre-mRNA:t innehåller otranslaterade regioner i båda ändar som innehåller ett ribosombindningsställe, terminator och start- och stoppkodon. Dessutom innehåller de flesta eukaryota oöversatta introner som tas bort innan exonerna översätts. Sekvenserna vid ändarna av intronerna dikterar splitsningsställena för att generera det slutliga mogna mRNA:t som kodar för proteinet eller RNA-produkten
Nedan visas strukturen för en eukaryot proteinkodande gen.
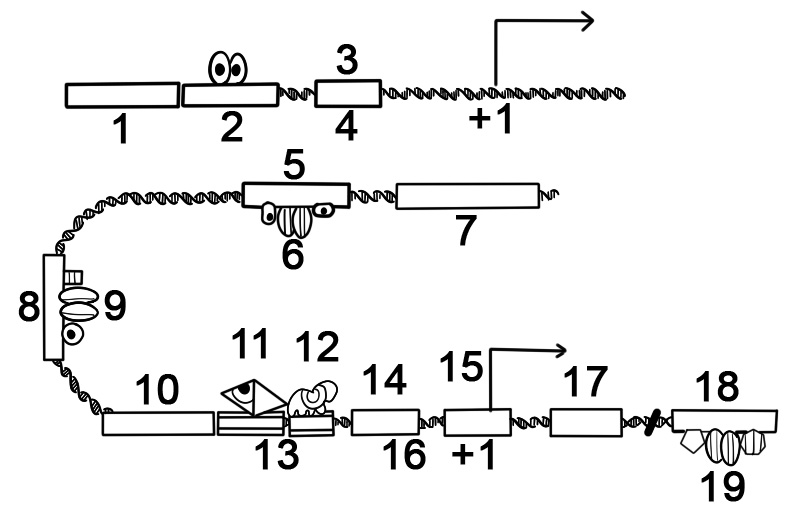
Många prokaryota gener är organiserade i operoner, med flera proteinkodande sekvenser som transkriberas som en enhet. Generna i en operon transkriberas som ett kontinuerligt mRNA, kallat polycistroniskt mRNA. I detta sammanhang är termen cistron ekvivalent med gen. Transkriptionen av en operons mRNA styrs ofta av en repressor som kan förekomma i ett aktivt eller inaktivt tillstånd beroende på närvaron av specifika metaboliter. När den är aktiv binder repressorn till en DNA-sekvens i början av operonet, kallad operatorregionen, och undertrycker transkription av operonet; när repressorn är inaktiv kan transkription av operonet inträffa. Produkterna av operongener har vanligtvis relaterade funktioner och är involverade i samma regulatoriska nätverk.
Nedan visas strukturen för ett prokaryot operon av proteinkodande gener.